分子シャペロン
(2009/05/16)
タンパク質のフォールディング
生命活動の万能の担い手であるタンパク質はアミノ酸が連なった「ひも」状のポリマーです。「ひも」というとぶらぶらの状態を想像するかもしれませんが、タンパク質はそのようなぶらぶらの状態では基本的にははたらくことはできません。アミノ酸の並び方(配列)に特有のある「かたち」(立体構造)に折れたたむ必要があるのです。その折れたたみはフォールディングと呼ばれていて、生命科学の重要な課題として長年研究が進められていますが、まだ十分に理解されているわけではありません。例えば、ヒトゲノムをはじめとする多くの生物のゲノム情報が解読されている現在、何万というタンパク質のアミノ酸配列情報はありますが、それらが最終的にどのような立体構造になるのかを知ることは今でも困難です。
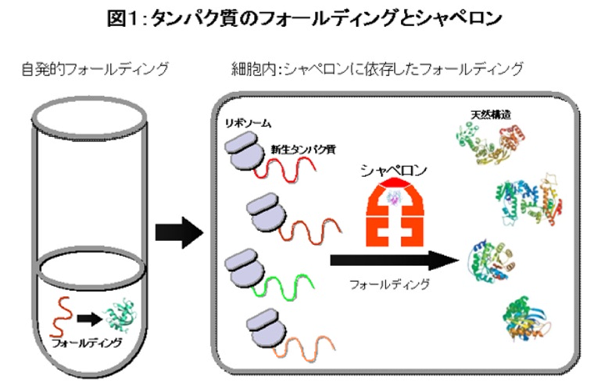
ただ、フォールディングというのは基本的には簡単でアミノ酸配列さえ決まれば自動的にフォールディングは完了する、ということが1950年代頃の試験管内でのフォールディング研究からわかっていました(図1)。言い方を換えると、タンパク質の立体構造はアミノ酸の「ひも」が最も安定な状態に落ち着くということです。その考え方は、その研究を行った人の名前からAnfinsenのドグマと呼ばれ、今でもタンパク質科学の基礎となっています。しかし、細胞の中でのフォールディングは一般に簡単ではありません。タンパク質がリボソームというタンパク質工場で産まれてきたばかりの「ひも」状態はとても不安定で、周りに同様の不安定な「ひも」があるとからまりあって凝集になってしまいます。また、フォールディングしたあともタンパク質は一般に不安定なので、高い温度にさらされたりするとかたちが崩れて(変性と言います)、凝集になります。卵をゆでるとゆで卵になるのが変性による凝集の例です(注:タンパク質は漢字で書くと蛋白質ですが、「蛋白」はもともと卵白という意味です)。
分子シャペロンとは
そうは言っても細胞内ではタンパク質が凝集など作らずにはたらいています。そこで登場するのが分子シャペロンです。分子シャペロンというタンパク質ファミリーは細胞内でタンパク質がきちんとフォールディングするのを助けています(図1)。分子シャペロンの元々の定義は、「他のタンパク質のフォールディングを助けるが、自らはその最終成分にならないタンパク質」ですが、現在では、タンパク質が産まれてから死に至るまで「タンパク質の一生」のさまざまな過程(膜透過、品質管理、タンパク質分解など)における介添えタンパク質全般を指す用語として使われています。(注:シャペロン(chaperone)の原義は、「デビューしたての若きレディーが(周辺の輩たちとの不適切な関係などを防ぎながら)一人前になるのを介添えする年配の婦人」というようなものです)
タンパク質をはじめとする生体高分子でたいへん混み合っている細胞内において、シャペロンはフォールディング途上の不安定な中間体や熱で変性したタンパク質が凝集にならないようにはたらきながら、フォールディングを助けています。代表的なシャペロンには、シャペロニンと呼ばれるファミリーがあり、ATPを使って多くのタンパク質のフォールディングを助けています。
日本生物物理学会における分子シャペロン研究
では、このシャペロニンはどのような仕組みでフォールディングを助けているのでしょうか。これまでに国内外で多くの研究が進められてきており、基本的な仕組みはわかってきました。ここでは日本生物物理学会のメンバーによって行われた研究成果を紹介いたします。
1分子レベルでのシャペロニン研究
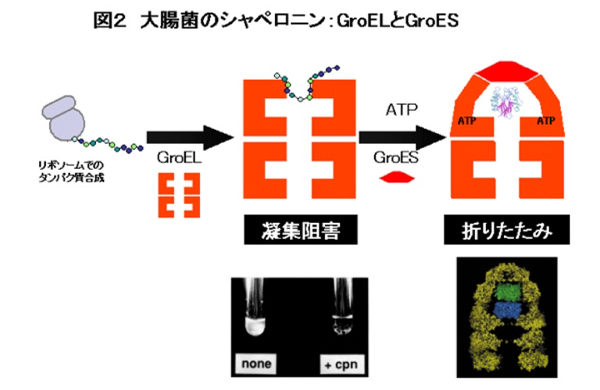
研究が最も進められてきているシャペロニンはバクテリアのGroELというタンパク質です。GroELは分子量が6万くらいのタンパク質が7つ筒のようになった直径約14nmのリングが二つ重なった構造をしています(図2)。GroELは変性タンパク質に結合して、凝集体形成を防ぐ能力を持ちます(注:図2の写真は好熱性バクテリアのGroELがあるときには卵白をゆでても固まらないようすを示しています)。GroELは補助因子としてはたらく「ふた」状のGroESとATPがあると複合体を形成します。そして、この複合体内にできる巨大な空洞の中で変性タンパク質のフォールディングが最後まで進行することがわかっています。その後、ATPが加水分解してADPになると、GroES、さらに空洞内のタンパク質も解離します(図2)。このGroELとGroESの結合と解離の仕組みはGroELの変異体を使ったりすることでわかってきたのですが、では、実際に直接ダイナミクスを観察して研究することはできないでしょうか。ここで、生物物理学的な手法が役に立ちました。蛍光1分子イメージングを応用して、GroELのはたらきを実時間で直接観察したのです。
(i) GroELとGroESの結合と解離の1分子観察:
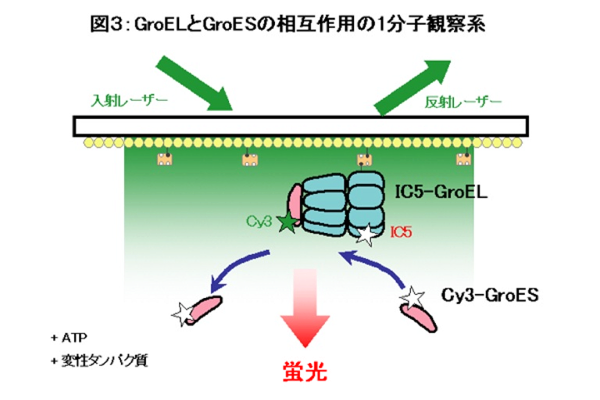
図3に示すような実験デザインで蛍光標識したGroELとGroESの結合・解離を観察しました。1分子の蛍光色素のイメージングには全反射蛍光顕微鏡によるエバネッセント照明を用いています。この顕微鏡は照射面から約150nmの領域しか励起しないため、背景光が劇的に減るだけでなく、スライドガラス表面に固定した蛍光標識タンパク質を局所励起できるという利点をもちます。まず、蛍光色素(IC5)で標識したGroELを固定して位置を決定しておき、別の蛍光色素(Cy3)で標識したGroESをATP存在下で加え、顕微鏡観察します。GroELに結合していないGroESはブラウン運動が激しいため蛍光スポットとなりませんが、GroESがGroELに結合すると固定によりブラウン運動が抑えられるために輝点として観察できます(動画でGroESの結合と解離のようすをご覧ください)。このような実験系で、単一のGroEL分子にGroESが結合、そして解離するようすがはじめて明らかとなりました。また、その結果を解析することにより、GroELとGroESの反応サイクルはそれまで知られていない中間体を経由して進むことが明らかとなりました。
(ii) 1分子蛍光イメージング技術を用いたGFPフォールディングの解析:
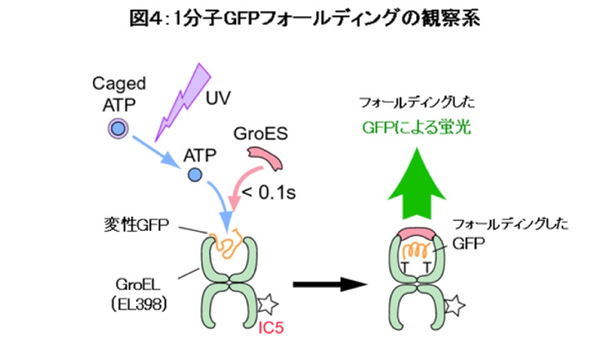
さらに、シャペロニン1分子内で起こるタンパク質のフォールディングを可視化する研究も行われています。シャペロニンの空洞内に閉じ込めるタンパク質としては、緑色蛍光タンパク質(GFP)を用います。GFPは変性状態では蛍光を発しませんが、フォールディングしたときだけ蛍光を発しますのでフォールディングしたかどうかを蛍光観察によりモニターすることができます。実験系としては、まず変性したGFPとGroELの複合体を作製してスライドガラスに固定し、その後GroES、caged ATPで観察セルを満たします(図4)。顕微鏡上で、この観察セルに紫外線を照射してcaged ATPからATPを生成したところ、GFP-GroEL複合体にGroESが結合してGFPのフォールディングが起こり、GFPの蛍光が観察できました。1分子レベルでタンパク質フォールディングを捉えることもできるのです。
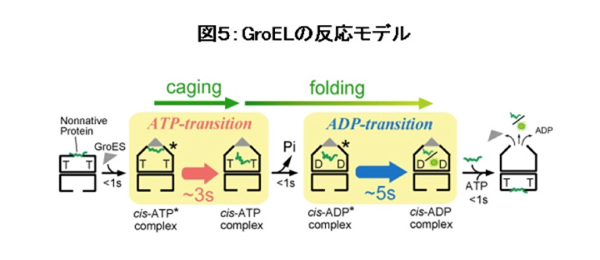
最後に、上記の1分子イメージングの解析結果なども含めて得られたGroELの反応サイクルモデルを図5に示します。GroELのような複雑なタンパク質の動態を1分子レベルで研究することにより、シャペロン研究に新たな方法論を導入し、さらには従来型の生化学研究に代表される多分子の平均からは得られない情報が得られました。
今後の課題 タンパク質のフォールディングの異常が病気に関わる例が多数知られています。アルツハイマー病、パーキンソン病、ハンチントン病、狂牛病(プリオン)など哺乳類の神経変性疾患に多く、深刻な問題となっているのはみなさんご存じの通りです。細胞内でのフォールディングに深く関わるシャペロンがこれらの疾患と関係していることも知られてきており、タンパク質のフォールディングやシャペロンの研究はますます重要になってきています。生物物理的な手法はここで示した1分子イメージング以外でもシャペロンやフォールディング研究の解明に多大な貢献をしています。また、生物物理を研究している人たちが有している学際的な研究スタイルがタンパク質科学の最重要課題の一つであるフォールディングの問題に新たな切り口を提示していきながら研究が進展していくものと思われます。
[参考文献]
Taguchi, H., Ueno, T., Tadakuma, H., Yoshida, M. & Funatsu, T. (2001) Single-molecule observation of protein-protein interactions in the chaperonin system. Nat. Biotechnol. 19, 861-865.
Ueno, T., Taguchi, H., Tadakuma, H., Yoshida, M. & Funatsu, T. (2004) GroEL mediates protein folding with a two successive timer mechanism. Mol Cell 14, 423-434.
東京工業大学・大学院生命理工学研究科 田口英樹(たぐちひでき)
https://www.taguchi.bio.titech.ac.jp/
東京大学・薬学研究科 船津高志