バクテリアべん毛モーター
(2010/05/24)
バクテリアの多くはべん毛と呼ばれる細長いらせん状繊維を菌体から伸ばし、それを根元のモーターで高速回転して推進力を発生し、水の中を活発に泳ぎ回ります。大腸菌やサルモネラ菌では左巻きらせんのべん毛が数本生えており、直線的に泳ぐ時はそれらが束になって推進力が発生します(図1)。モーターが数秒に一度急反転すると、そのねじれ力によって瞬時にらせんが右巻きに変換し、その結果べん毛の束がほぐれて推力のアンバランスを生じ、菌体が回転して遊泳方向を転換します。モーターの回転方向の切り替えは、細胞膜に存在する化学温度センサーが発信する信号によって制御され、その結果細菌は栄養や温度などが最適な環境に集まることができます。ここでは、べん毛の自己構築機構および回転機構について概説します。
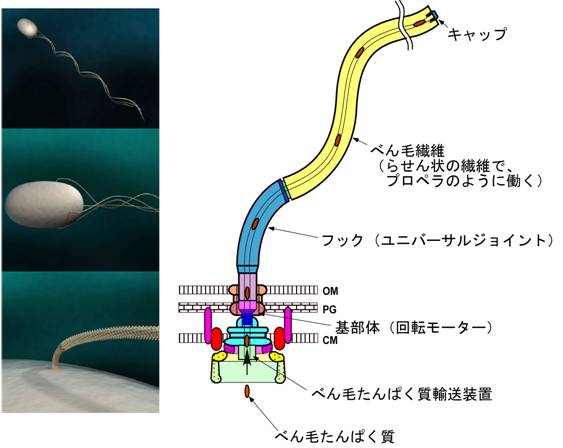
図1. べん毛で泳ぐ細菌の模式図と細菌べん毛の模式図
サルモネラ菌や大腸菌は数本のべん毛を細胞周辺から延ばし、束にして回転させて泳ぎます。べん毛は大まかに回転モーターである基部体、ユニバーサルジョイントであるフック、プロペラのように動くべん毛繊維の、3つの部分構造で構成されています。(CM;細胞膜/PG;ペプチドグリカン層/OM;外膜)
(1)べん毛構造 べん毛は、約30種類の蛋白質からなる超分子複合体で、基部体、フック、繊維と呼ばれる、おおまかに3つの部分構造から構成されます(図1)。基部体は細胞表層膜系内に存在し、Cリング、MSリング、 Pリング、Lリング、およびロッドと呼ばれる軸構造からなり、回転モーターとして働きます。繊維はフラジェリンと呼ばれる1種類の蛋白質が2~3万分子重合した長さ十数 μm の細長いらせん状繊維で、分子プロペラとして機能します。曲げには柔らかいが、ねじれには硬い性質をもつフックは、フック蛋白質が約120分子らせん状に重合したチューブ状構造で、基部体の発生する回転力を繊維に伝えるユニバーサルジョイントとして働きます。
(2)べん毛構築過程
べん毛は基部体、フック、繊維の順に構築されます(図2)。まずは、FliFが細胞膜内で自己集合してMSリングを形成します。次に、FliG、FliM、およびFliNがMSリングの細胞質側に自己集合し、Cリングを形成します。MS-Cリング複合体が完成すると、細胞膜を越えて存在するロッド-フック-繊維と連なるべん毛軸構造体の構築が開始します。これらべん毛軸構成蛋白質は、べん毛の基部に存在する独自の蛋白質輸送装置によって認識され、べん毛軸構造体の中心を貫通する直径2 nmの細長いチャネルの中へ、そしてべん毛先端へと輸送されます。最初に、ロッドが作られます。ロッドができるとその周りにLPリング複合体が形成され、基部体が完成します。次に、ロッド先端にフックの形成を助けるフックキャップができると、その直下に新たに送り出されてきたフック蛋白質が重合します。フックの長さが約55 nmに達すると、フック-フィラメント連結蛋白質が送り出されます。その結果、フックキャップがフックの先端から外れ、フック-フィラメント連結部が作られます。この連結部の先端に繊維の構築を助けるフィラメントキャップができると、その直下にフラジェリンが重合し、繊維を形成します。
べん毛の形成には約40個のべん毛遺伝子が必要です。これらの遺伝子の発現はランダムに起こるのではなく、べん毛の形成過程に応じて制御されます。フックが完成するまでは転写抑制因子が細胞内に蓄積し、その結果べん毛繊維の形成に必須な遺伝子群の発現は抑制されます。フックが完成すると、転写抑制因子が輸送装置によって細胞外に排出され、その結果それらの遺伝子群の発現が誘導され、べん毛繊維が構築されます。このように、べん毛基部体の細胞質側にある輸送装置は遙かに離れたフック先端で起こるフックの重合過程をモニターし, その情報にもとづいて輸送すべき蛋白質を配送する仕組みを備えています。このような輸送装置の選択的物質輸送機能により、べん毛は効率的かつ順序正しく構築されます。興味深いことに、フラジェリンの高発現は細胞にとって大きな負荷となり、著しい増殖阻害が引き起こされることから、このような蛋白質輸送の制御システムは細胞のエネルギー効率にとっても大切と言えます。

図2. べん毛の構築モデル。
約30種類の異なった蛋白質が順序通りに組み込まれ、構築される。左上から右下へ進む。
(3)べん毛モーターの回転機構
大腸菌やサルモネラ菌のべん毛モーターが回転するためのエネルギー源は、細胞膜内外に形成されるプロトンの電気化学的ポテンシャル差、すなわちプロトン駆動力です。一方、海洋性ビブリオ菌の極べん毛モーターや強いアルカリ性環境でのみ生育できる好アルカリ細菌のべん毛モーターは、ナトリウム駆動力で回転します。興味深いことに、枯草菌のべん毛モーターは、pH、ナトリウムイオン濃度などの環境の変化に応じてプロトン駆動力とナトリウム駆動力を使い分けます。
大腸菌およびサルモネラ菌のべん毛モーターのトルク発生には、MotA、MotB、FliG、FliM、およびFliNの5種類の蛋白質が関与します(図3)。MotAとMotBは細胞膜貫通型の蛋白質で、4分子のMotAと2分子のMotBからなる複合体を形成し、プロトン透過経路として働きます。べん毛基部体の周りには、少なくとも11個のMotA/B複合体が存在します。MotBのC末ペリプラズムドメイン(MotBC)にはペプチドグリカン結合モチーフと呼ばれるアミノ酸配列が存在することから、MotA/B複合体がべん毛モーターの固定子として機能すると考えられています。最近、X線結晶構造解析によってピロリ菌およびサルモネラ菌のMotBCの立体構造が明らかにされ、その構造がPalやRamと言ったペプチドグリカン結合蛋白質と良く似た構造であることがわかりました。興味深いことに、GFP-MotBの一分子蛍光イメージングにより、MotA/B複合体は常にペプチドグリカン層に結合しているのでなく、モーター回転中にもダイナミックに入れ替わることが示されました。FliG、FliM、およびFliNはスイッチ蛋白質とも呼ばれ、MSリング直下にCリングと呼ばれるカップ状の構造を形成し、べん毛モーターの回転子として機能します。これらのスイッチ蛋白質は、べん毛モーターの回転方向の切り替えスイッチとしても働きます。走化性応答においてリン酸化されたCheYがFliMおよびFliNに結合すると、べん毛モーターの回転方向が切り替わります。
プロトンがMotA/B複合体を流れると、MotA の細胞質ドメインがFliGと相互作用し、その結果べん毛モーターの回転力が発生すると考えられています。大腸菌内で動作するように作られたナトリウムイオン駆動型モーターを用いて回転計測を行った結果、べん毛モーターが26回のステップによって1回転することが明らかされました。この結果は、回転子であるFliGが作るリングの対称性が26であることと一致していました。
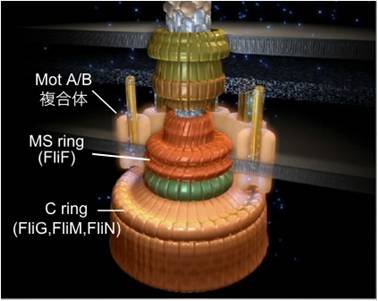
図3. べん毛モーターの模式図
べん毛モーターのトルク発生には、MotA、MotB、FliG、FliM、およびFliNの5種類の蛋白質が関与する。MotAおよびMotBはべん毛モーターの固定子複合体を形成する。FliG、FliM、およびFliNはMSリングの細胞質側に自己集合してCリングを形成する。
[参考文献]
[1] Berg, H.C. (2003) The rotary motor of bacterial flagella. Annu Rev Biochem 72: 19-54.
[2] Minamino, T., Imada, K., Namba, K. (2008) Mechanisms of type III protein export for bacterial flagellar assembly. Mol Biosyst 4: 1105-15
[3] Minamino, T., Imada, K., Namba, K. (2008) Molecular motors of the bacterial flagella. Curr Opin Struct Biol 18: 693-701.
[4] Sowa, Y., Berry, R.M. (2008) Bacterial flagellar motor. Q Rev Biophys 41: 103-132.
大阪大学・大学院生命機能研究科,JST・さきがけ研究員 南野 徹